雷公藤红素,又名南蛇藤素,是从雷神藤中分离出的最具前途的天然药用产物之一,对多种肿瘤类型均有较显著的抑制作用。然而,雷公藤红素具有较严重的副作用,所以它的临床应用受到严格限制,同时,由于其发挥抗肿瘤活性的分子靶标和作用机制不够清晰,缺乏雷公藤红素与靶标的复合物晶体结构,使得基于其化学结构改造开发高效低毒的雷公藤红素衍生物缺乏明确的指导方向。
2023年2月3日,中科院上海药物所张豪与上海交通大学徐颖、张翱为共同通讯作者在《Signal Transduction and Targeted Therapy》(IF=38.104)发表了题为:“Celastrol suppresses colorectal cancer via covalent targeting peroxiredoxin 1”的研究论文,在这里,作者开发了一种新的靶标富集排序工具—OTTER,并利用这个新工具结合生物化学、生物物理等技术手段,发现过氧化物还原酶1(peroxiredoxin 1, PRDX1)是结直肠癌中雷公藤红素的靶蛋白,并首次解析了雷公藤红素与PRDX1蛋白的复合物晶体结构,又基于晶体结构分析,设计并合成了特异性及体内安全性显著提高的雷公藤红素衍生物19-048,证明了雷公藤红素的体内毒性,可以通过靶标发现及基于结构的化学优化而降低,同时表明,抑制PRDX1有望用于治疗结直肠癌。值得注意的是,本研究使用了汉恒生物提供的针对PRDX1的CRISPR-Cas9慢病毒。
下面,我们一起来看具体的研究结果:
首先,作者通过RNA测序得到用雷公藤红素处理后的差异表达基因,然后用OTTER富集雷公藤红素的氧化还原相关靶蛋白,发现雷公藤红素对过氧化物还原酶(peroxiredoxin, PRDX)蛋白家族的若干成员有较明显的酶活抑制,其中,雷公藤红素对PRDX1蛋白的酶活抑制最强,进一步的质谱数据以及点击化学实验提示,雷公藤红素与PRDX1的第173位半胱氨酸共价结合。
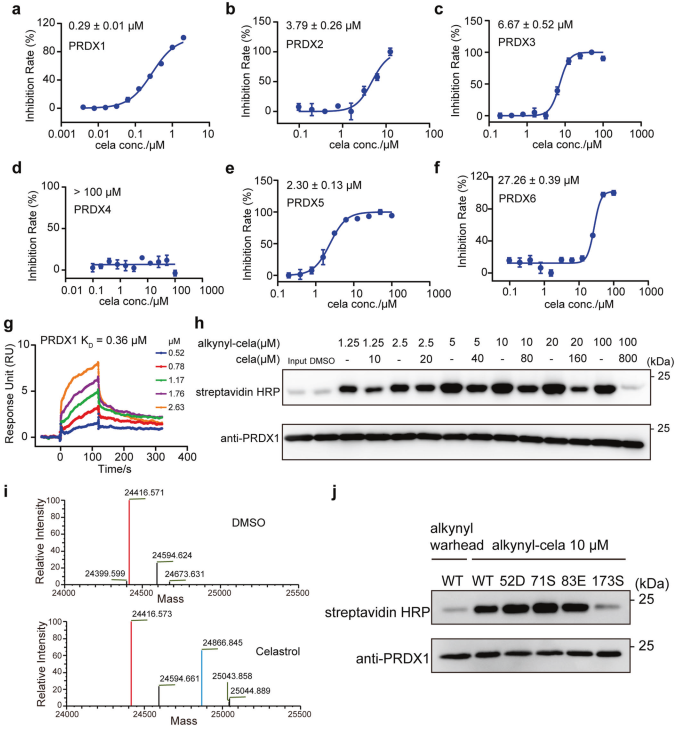
图1.雷公藤红素抑制PRDX1酶活
先前的研究预测,雷公藤红素以立体特异性方式与靶蛋白共价结合,这可能会影响雷公藤红素的靶蛋白选择性,作者将PRDX1与雷公藤红素复合物晶体结构解析为1.76埃,揭示了其独特的结合模式。每个PRDX1蛋白二聚体,能够共价结合两分子的雷公藤红素,并且这两分子雷公藤红素能够与临近的PRDX1蛋白二聚体形成非共价的氢键以及疏水相互作用。
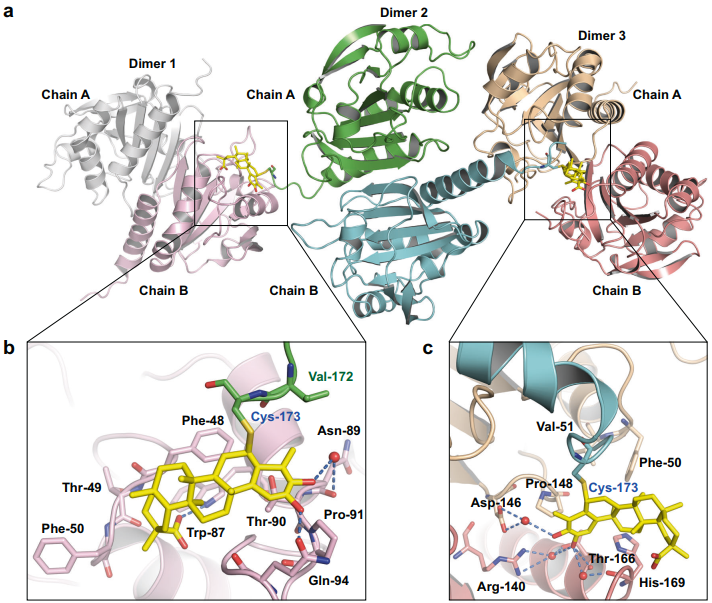
图2.雷公藤红素与PRDX1蛋白的复合物晶体结构
为了获得更高效和更具有特异性的新型PRDX1,作者合成了雷公藤红素的衍生化合物,其中19-266与19-048通过胍末端取代法合成,而19-048对PRDX1表现出高效的特异性和酶活抑制效果。
由于PRDX1通过清除ROS密切参与人类结直肠癌的发展,所以作者又证实了雷公藤红素和19-048治疗后能抑制结直肠癌细胞的增殖以及降低细胞活力。为了确定雷公藤红素和19-048是否通过靶向PRDX1影响细胞氧化还原状态,作者在SW620和HCT116细胞中用DCFH-DA荧光探针染色检测ROS水平,在不同浓度的雷公藤红素和19-048处理后,与对照相比,细胞中的ROS含量急剧升高,随后检测了雷公藤红素和19-048对NCM460细胞中ROS水平的影响,结果表明,雷公藤红素和19-048对正常细胞ROS诱导的影响小于癌细胞,后面作者又证明了雷公藤红素和19-048诱导的ROS升高会导致结直肠癌细胞的周期停滞和凋亡。
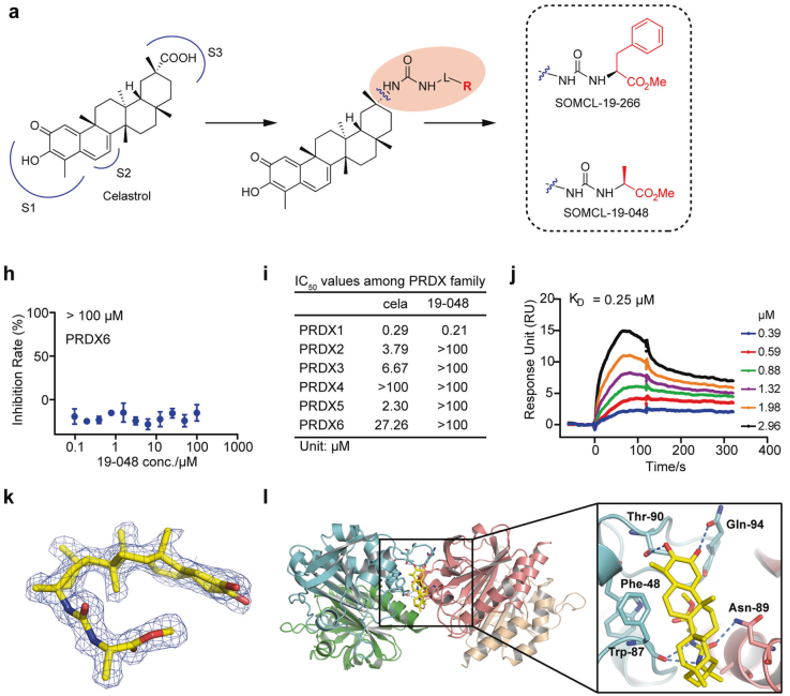
图3.雷公藤红素衍生物19-048具有更高的效力和特异性
为了验证PRDX1是雷公藤红素和19-048在结直肠癌细胞中诱导细胞周期停滞和凋亡的主要靶蛋白,作者在SW620细胞中通过CRISPR-Cas9系统构建了PRDX1敲低细胞系(SW620-gPRDX1),用阴性sgRNA(SW620-gNS)作为对照,与对照相比,SW620-gPRDX1细胞中ROS积累增加,用雷公藤红素或19-048处理进一步增强了ROS升高的程度,相反,PRDX1过表达显著逆转了雷公藤红素和19-048诱导的ROS升高。而且观察到SW620-gPRDX1的G1/M细胞周期停滞和细胞凋亡增强,过表达PRDX1显著减弱了雷公藤红素和19-048诱导的这些作用。这些结果表明,PRDX1是雷公藤红素和19-048在结直肠癌细胞中的主要靶蛋白,PRDX1的抑制导致细胞内ROS升高以及细胞周期停滞和凋亡增加。
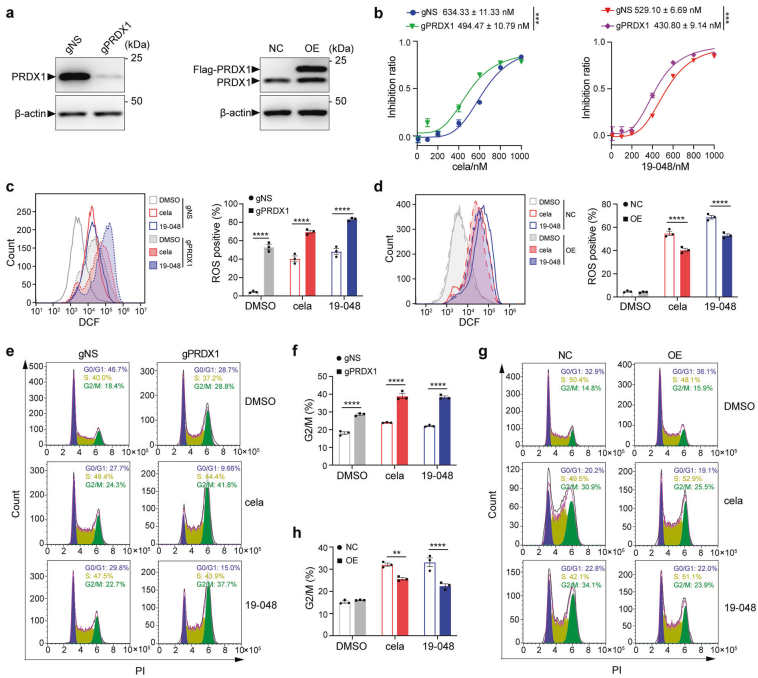
图4.雷公藤红素和化生物19-048通过靶向PRDX1抑制结直肠癌细胞增殖
为了在转录组水平上证明雷公藤红素和化合物19-048在结直肠癌细胞中的作用,作者收集并分析了SW620和HCT116细胞的RNA测序数据,然后用雷公藤红素和19-048处理细胞后获得差异表达基因(DEG),使用生物信息学方法分析显示,P53信号通路是其中排名最高的癌症相关信号通路。根据RNA测序数据,作者分别从SW620和HCT116细胞中总共选出了来自p53通路中的26个DEG,并用雷公藤红素和19-048处理后测定它们的RNA表达水平。QPCR和WB结果表明,用雷公藤红素和19-048处理后,这些DEG的表达水平升高,而ROS清除剂-NAC阻断了这些基因的转录上调。总之,这些结果表明,在雷公藤红素和化合物19-048的存在下,激活的p53下游基因参与了PRDX1抑制引起的细胞周期停滞和凋亡。
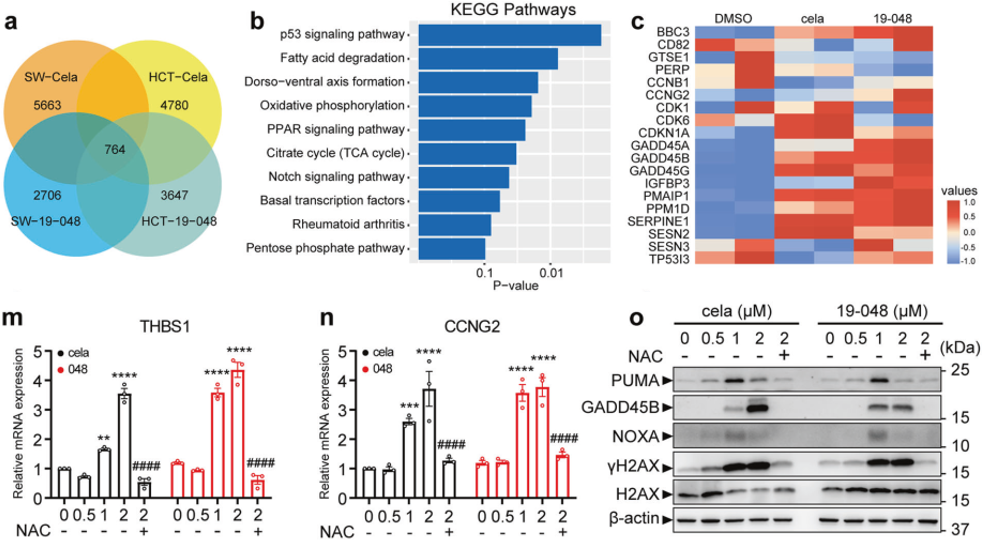
图5.雷公藤红素和19-048影响的相关信号通路和基因
为了确定PRDX1是否是雷公藤红素和19-048在体内的主要靶蛋白,作者使用已建立的SW620-gPRDX1和SW620-gNS细胞进行裸鼠成瘤,来评估PRDX1敲低对这些化合物抗肿瘤功效的影响。当肿瘤生长到50mm时,小鼠用雷公藤红素和19-048进行治疗,结果表明雷公藤红素和19-048抑制了SW620-gNS肿瘤的进展。同时,作者对小鼠进行了毒性试验,结果表明,与雷公藤红素相比,19-048在体内的安全性显著提高。免疫组织化学染色显示,在SW620-gNS移植小鼠中,随着雷公藤红素和19-048处理,GADD45G和γ-H2AX蛋白水平升高,而PRDX1的敲低减弱了雷公藤红素和19-048导致的GADD45G和γ-H2AX蛋白的上调,与肿瘤体积和重量的趋势一致。这些结果表明,雷公藤红素和化合物19-048通过靶向PRDX1抑制体内结直肠癌肿瘤生长。

图6.雷公藤红素和19-048通过靶向PRDX1抑制结直肠癌的肿瘤生长
综上,作者用他们新开发的靶标富集排序工具,结合生化以及生物信息学手段,发现雷公藤红素的靶标-PRDX1,通过雷公藤红素与PRDX1复合物的晶体结构,建立了雷公藤红素如何与靶标蛋白发生共价修饰作用的结构基础,同时,基于晶体结构分析,作者设计合成的雷公藤红素的衍生物19-048表明雷公藤红素的副作用可以通过结构修饰来减少。此研究为更多重要天然产物的靶标发现提供了可以借鉴的研究策略以及实用的靶标发现新工具。









 3344
3344










