前几期干货为大家介绍了组织特异性表达调控,其中涉及到了重组酶系统,本期小恒将为大家介绍“Cre/Loxp及其相关重组酶系统”。

Cre-LoxP系统是一种重组酶系统,能在DNA特定位点上执行删除、插入、易位及倒位,是一种广泛应用的基因编辑工具,可以达到在基因水平上对生物体进行定向遗传改造的目的,在真核和原核系统中均适用。
Cre(Cyclization Recombination Enzyme)是一种重组酶,1981年从P1噬菌体中发现,基因编码区序列全长1029bp,编码由343个氨基酸组成的38kDa单体蛋白。Cre重组酶不仅具有催化活性,而且与限制酶相似,能识别特异的DNA序列,即Loxp位点,介导Loxp位点间的基因序列删除或重组。
LoxP(locus of X-over P1)来源于噬菌体P1,是一段长度为34bp的DNA序列,由两个13bp的反向重复序列和一个不对称的8bp间隔区共同组成。反向重复序列是Cre重组酶的特异识别和结合区域,而间隔区域决定了LoxP位点的方向。
其序列如下:(N 表示碱基可能发生变化)
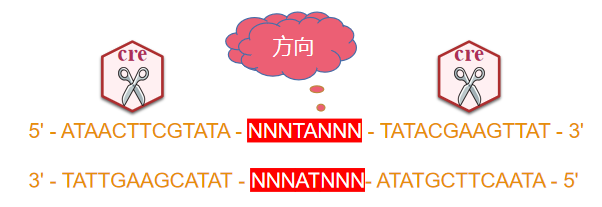
图1 Cre-LoxP基本作用原理
一、Cre-LoxP系统基本作用方式
一般而言,当细胞基因组内存在两个LoxP位点时,Cre重组酶会诱导两个LoxP位点间的序列发生重组。重组的结果取决于两个LoxP位点的方向,主要有以下几种可能:
1、如果两个LoxP位点位于一条DNA链上,但方向相反,Cre重组酶能导致两个LoxP位点间的序列翻转;
2、如果两个LoxP位点位于一条DNA链上,并且方向相同,Cre重组酶能有效切除两个LoxP位点间的序列;
3、如果两个LoxP位点分别位于两条不同的DNA链或染色体上,Cre酶能介导两条DNA链的交换或染色体易位;
4、如果四个LoxP位点分别位于两条不同的DNA链或染色体上,Cre重组酶能诱导LoxP间的序列互换。
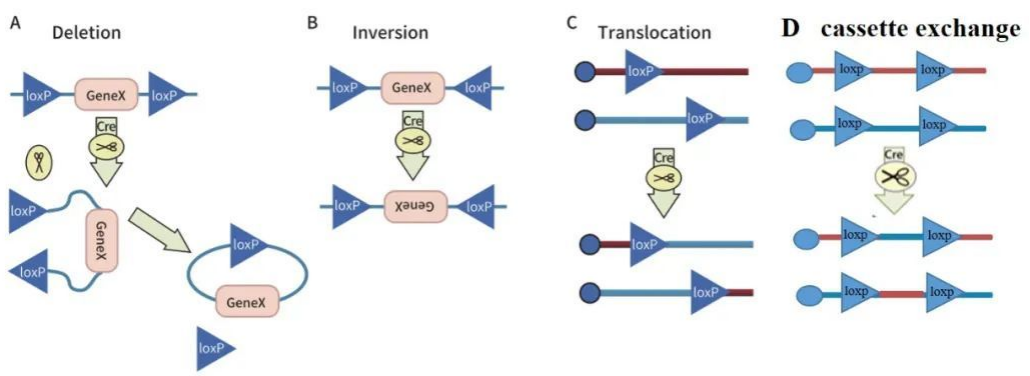
图2 Cre-LoxP诱导基因重组的方式
二、Cre-LoxP条件性重组改造
经过现代基因工程方法对Cre和LoxP元件的改造,Cre-LoxP系统实现了更加丰富的条件性重组策略。
1、Cre元件改造
在Cre元件的C端接上一段改造过的配体结合结构域LBD,新的融合蛋白Cre-LBD将定位在胞浆内,当人工合成的激素分子结合到Cre-LBD受体后,蛋白构象改变并进入核内,介导基因重组,目前使用最多的是Tamoxifen诱导的CreERT2突变体,它的LBD来自于雌激素受体ER分子,当有Tmoxifen的时候,Cre才能介导基因重组,这样通过控制Tamoxifen的注射时间,可以实现对基因重组时间特异性的调控。
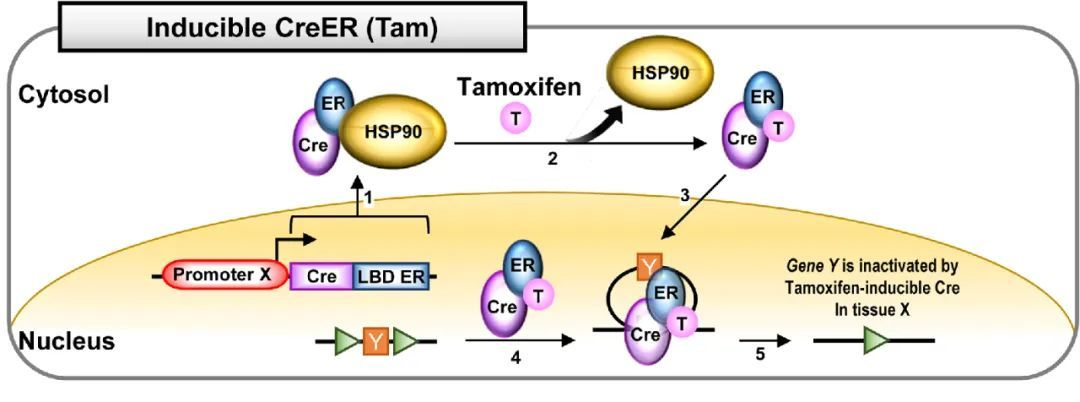
图3 Tamoxifen诱导的Cre-ER系统
2、Loxp元件的改造
LoxP元件也有一些突变体,间隔区和回文序列都可以进行突变,突变后的序列依然能被Cre重组酶识别和重组,但是突变的LoxP序列必须和同样突变的LoxP序列匹配介导基因重组,而不能和未突变的LoxP序列匹配,这样将不同的LoxP序列组合用于控制多个基因(DIO/DO系统中的Lox2272位点),在同一Cre重组酶的作用下,可以实现多序列的基因重组,产生非常多元的重组结果。
DIO/DO序列
通过引入两对不同的LoxP位点—LoxP和Lox2272,经过两组Lox位点的两轮重组可达到一种稳定状态。也就是可以通过Cre重组酶的存在与否来控制基因的表达。
在这种策略下,任一对 Lox 位点间的序列会在 Cre 的作用下发生可逆的快速翻转,之后 Cre 重组酶马上不可逆地切割翻转后的同向 Lox 位点,只留下翻转后的基因和单独的 LoxP、Lox2272 位点,防止基因的再次翻转。基于上述原理,可以在病毒载体中构建 DIO 和反向基因,感染 Cre 阳性细胞后,可以让Cre阳性的细胞表达基因,称为DIO(Cre-on) 结构;如果预先包装的基因是正向,那么 Cre 阳性的细胞则不表达基因,其余细胞表达基因,称为DO(Cre-off)结构。
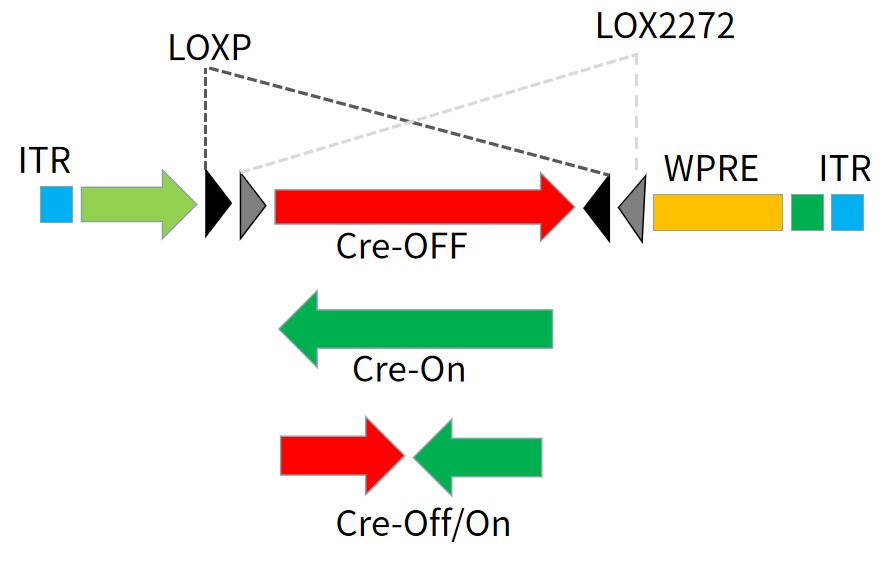
图4 借助LoxP和Lox2272的DIO/DO策略实现Cre依赖的基因表达
三、其他重组酶系统介绍
重组酶系统除了经典的Cre-Loxp系统外,还有与Cre-Loxp系统无交叉影响的vCre-vLoxP、sCre-sLoxP 系统、Flp/Flpo-FRT系统、Dre-Rox 系统和Vika-vox系统。
1、vCre/sCre系统
vCre/sCre均为Cre重组酶的同源蛋白,但与Cre的同源性很低,可以特异性识别vLoxp/sLoxp位点。
2、Flp-FRT系统
Flp(Flippase recombination enzyme)重组酶,从酵母细胞内被发现,其基因全长1272bp,为48kDa大小的、由423个氨基酸组成的多肽单体蛋白。FRT是Flp的识别位点,全长48bp,包含3个13 bp的重组酶结合区和1个8 bp的含Xba I位点的非对称间隔区(spacer)。Flp-FRT 系统与 Cre-LoxP 诱导基因重组的方式类似,两系统明显的区别是重组酶(Cre 和 Flp)具有不同的最佳反应温度,有研究发现,Cre 重组酶的最佳温度为 37℃,而 Flp 重组酶为 30℃。研究人员后续构建了Flpo—Flp突变体,在37℃也可以表现出较好的耐热性。
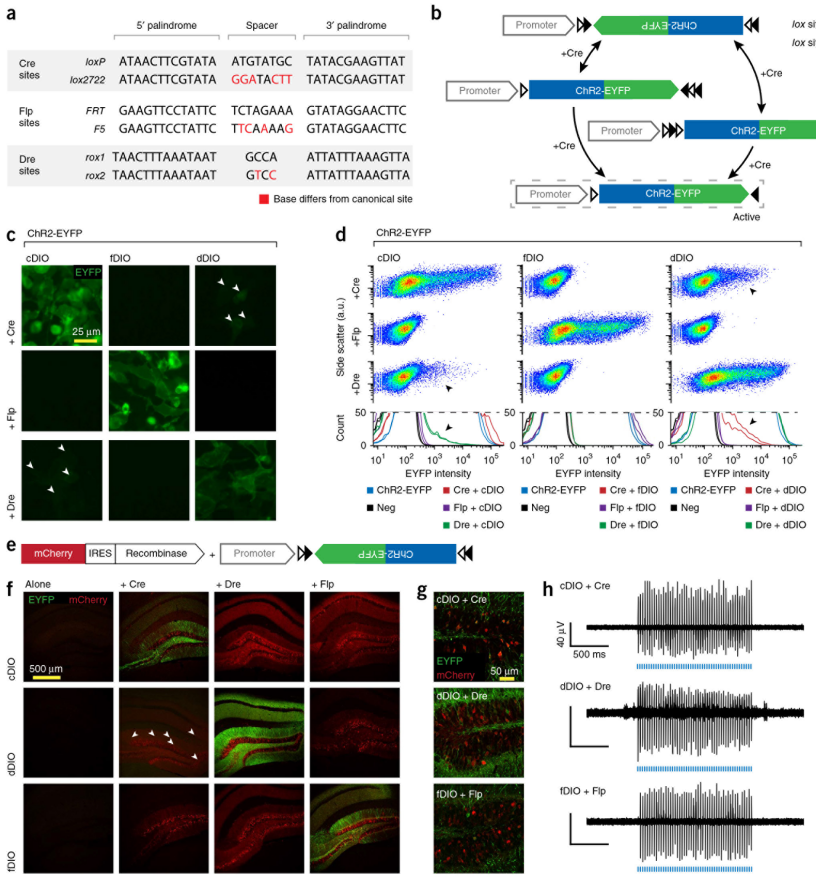
图5 Cre\Flp\Dre 系统比较
3、Dre-Rox系统
Dre重组酶来源于D6噬菌体,能特异性地识别一段长度为32bp的DNA序列(Rox位点)并介导该序列间的DNA发生重组。因其重组效率高且不会与Cre重组酶发生交叉反应,二者常联合使用以实现更加精细的基因操作。
4、Vika-vox系统
Vika重组酶来源于珊瑚弧菌,基因全长1086bp,能特异性地识别一段长度为34bp的DNA序列(vox位点)并介导该序列间的DNA发生重组。该系统与Cre/Loxp和Flp/FRT系统相比,具有细胞毒性低,使用范围广和安全性高等优势。
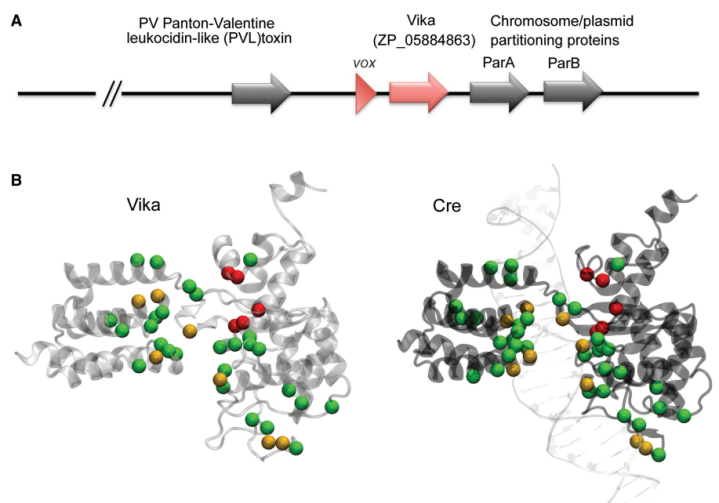
图6 Vika 及 Cre结构模型
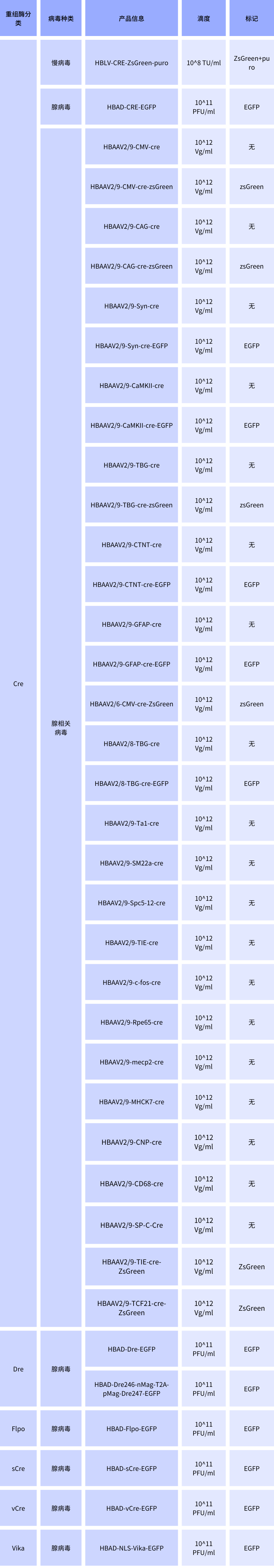
四、汉恒重组酶病毒现货列表
五、Cre-Loxp系统的应用案例
1、依赖Cre的条件性基因表达
文章利用AAV血清型和特异性启动子结合Cre-Loxp系统在Cre小鼠特定组织实现目的基因的表达;如:将AAV5-hSyn-DIO-hM3D(Gq)-mCherry通过脑立体定位注射到Vglut3-Cre小鼠脑DRN区,结合化学遗传实现DRN区神经元细胞的激活。
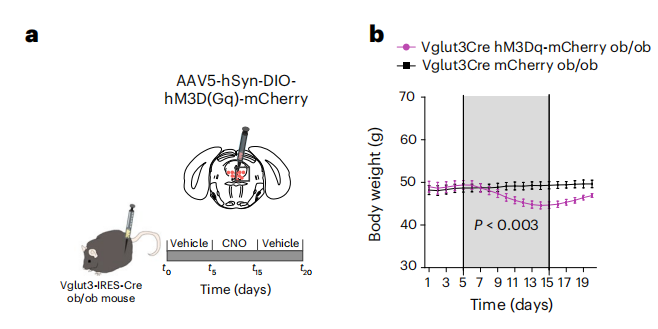
图7 依赖Cre介导的基因条件性表达
2、依赖Cre的条件性基因敲除
文章利用AD介导的Cre-Loxp系统可以对特定组织实现目的基因的快速敲除。如:利用AD载体携带Cre酶基因,注射到PINCH-1flfl和KrasLSL-G12D/+(Krasflfl/+)工具鼠注入肺诱导KrasG12D的表达和PINCH-1基因的失活。PINCH-1敲除显著降低了细胞增殖,KrasLSL-G12D/+小鼠表达KrasG12D显著诱导肺肿瘤形成。
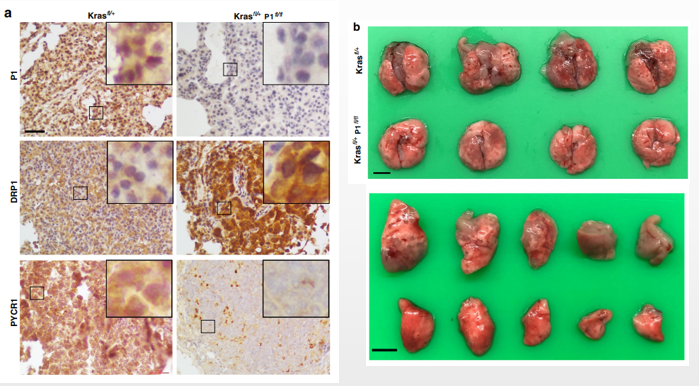
图8 依赖Cre介导的特定组织基因敲除
3、依赖cre的条件性基因敲低
文章利用AAV-shRNA通过脑立体定位注射Crh-IRES-Cre小鼠PVN区,在PVN区实现PSD-93基因的敲低。
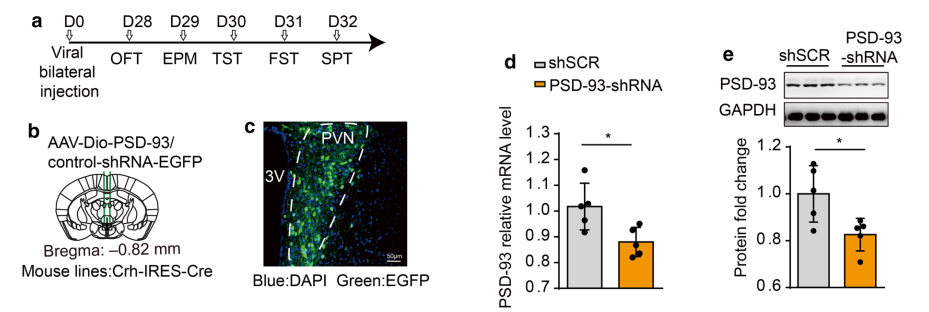
图9 依赖Cre介导的特定组织基因敲低
六、Cre-LoxP系统的优点
Cre-LoxP系统是目前在神经系统中应用最广泛的条件性基因敲除工具,主要是因为该系统有如下优点:
1、高效性:Cre重组酶与具有LoxP位点的DNA片段形成复合物之后,可以提供足够的能力引发之后的DNA重组过程,重组过程简约高效;
2、特异性强:LoxP位点是一段含回文序列结构和中间有间隔的34bp元件,这种结构保证了LoxP序列的唯一性,从而保证基因重组的特异性很强;
3、应用范围广:Cre重组酶是一种比较稳定的蛋白质,可以在生物体不同的组织、不同的生理条件下发挥作用;
4、II型启动子启动表达:Cre重组酶的编码基因可由任何一种II型启动子驱动,由此保证Cre重组酶在生物体不同的细胞、组织、器官或者在不同的发育阶段或不同的胜利条件下表达,从而实现较高的组织和细胞特异性。
至此,本期内容就结束了,主要介绍了“Cre/Loxp及其相关重组酶系统”,下期我们将分享心血管系统特异性表达调控,敬请期待。
参考文献:
[1] Kim, H., Yonsei University College of Medicine, Seoul, Kim, M., Yonsei University College of Medicine, Seoul, Im, S.K., Yonsei University College of Medicine, Seoul, et al. Mouse Cre-LoxP system: general principles to determine tissue-specific roles of target genes[J]. Laboratory Animal Research, 2018.
[2] Schneeberger M, Brice NL, Pellegrino K, Parolari L, Shaked JT, Page KJ, Marchildon F, Barrows DW, Carroll TS, Topilko T, Mulligan VM, Newman R, Doyle K, Bürli R, Barker DF, Glen A, Ortuño MJ, Nectow AR, Renier N, Cohen P, Carlton M, Heintz N, Friedman JM. Pharmacological targeting of glutamatergic neurons within the brainstem for weight reduction. Nat Metab. 2022 Nov;4(11):1495-1513.
[3] Ling G , Cui C . PINCH-1 regulates mitochondrial dynamics to promote proline synthesis and tumor growth[J]. Nature Communications, 2020, 11(1):4913.
[4] Qin XY, Shan QH, Fang H, Wang Y, Chen P, Xiong ZQ, Swaab DF, Zhou JN. PSD-93 up-regulates the synaptic activity of corticotropin-releasing hormone neurons in the paraventricular nucleus in depression. Acta Neuropathol. 2021 Dec;142(6):1045-1064.
[5] Fenno L E , Mattis J , Ramakrishnan C , et al. Targeting cells with single vectors using multiple-feature Boolean logic[J]. NATURE METHODS, 2014, 11(7):763-772.
[6] Madina K , Josephine A G , Nicolas B , et al. Vika/vox, a novel efficient and specific Cre/loxP-like site-specific recombination system[J]. Nucleic Acids Research(2):e37-e37.









 7651
7651










