内质网(ER)应激与多种代谢疾病有关,包括代谢功能障碍相关的脂肪变性肝病(MASLD)。内质网膜蛋白复合物(EMC)在内质网应激、膜蛋白转运异常和内质网降解等细胞过程中起调节作用。EMC10是内质网膜蛋白复合物的一个亚基,包括两种亚型,在调节能量稳态和肥胖中发挥作用,而这两种亚型在MASLD中的作用尚未区分。

2024年4月7日,复旦大学王宣春团队在Journal of Hepatology(IF=26)在线发表题为“EMC10 modulates hepatic ER stress and steatosis in an isoform specific manner”的研究论文,该研究揭示了EMC10以同种异构体特异性方式调节肝脏ER应激和脂肪变性,分泌型亚型(scEMC10)促进肝细胞PERK-eIF2α-ATF4信号的激活,而ER膜结合型亚型(mEMC10)抑制这些信号的激活;在接受MCD饮食或衣霉素(tunicamycin)处理的小鼠中,EMC10基因敲除加重了肝脏ER应激和脂肪变性,而过表达mEMC10改善了这种情况;并且作者通过抗体中和作用将scEMC10确定为MASLD的一种易治疗靶点。汉恒生物有幸为作者提供了腺相关病毒AAV8-scEMC10和AAV8-mEMC10,以实现这两种亚型的在体表达。
EMC10消融改善HFD喂养小鼠的肝脂肪变性
作者在之前的研究中已经证明EMC10消融降低了高脂肪饮食(HFD)喂养小鼠的肝脏脂质含量。为了进一步探索EMC10在肝脂肪变性中的作用,作者对HFD饲喂的EMC10 KO(Emc10 knockout)小鼠肝脏健康进行了表型评估。结果显示,在HFD饲喂条件下,Emc10 KO小鼠比野生型(WT)更瘦,但肝脏指数(肝脏质量与体重之比)没有差异。与WT相比,KO小鼠的肝脂肪变性明显减少,肝脂肪含量(LFC)、血浆脂质和谷草转氨酶(AST)水平显著降低;参与脂质代谢的三类基因:脂肪生成、脂肪酸(FA)摄取和FA β氧化相关基因的mRNA水平显著低于WT小鼠。肝脂肪变性常伴有肝脏炎症和纤维化,与WT相比,KO小鼠肝脏的炎症得到了很大改善,而纤维化保持不变。与ER应激相关的一些基因,例如ATF4和CHOP的表达显著降低。这说明EMC10的消融可以预防HFD诱导的肝脂肪变性。
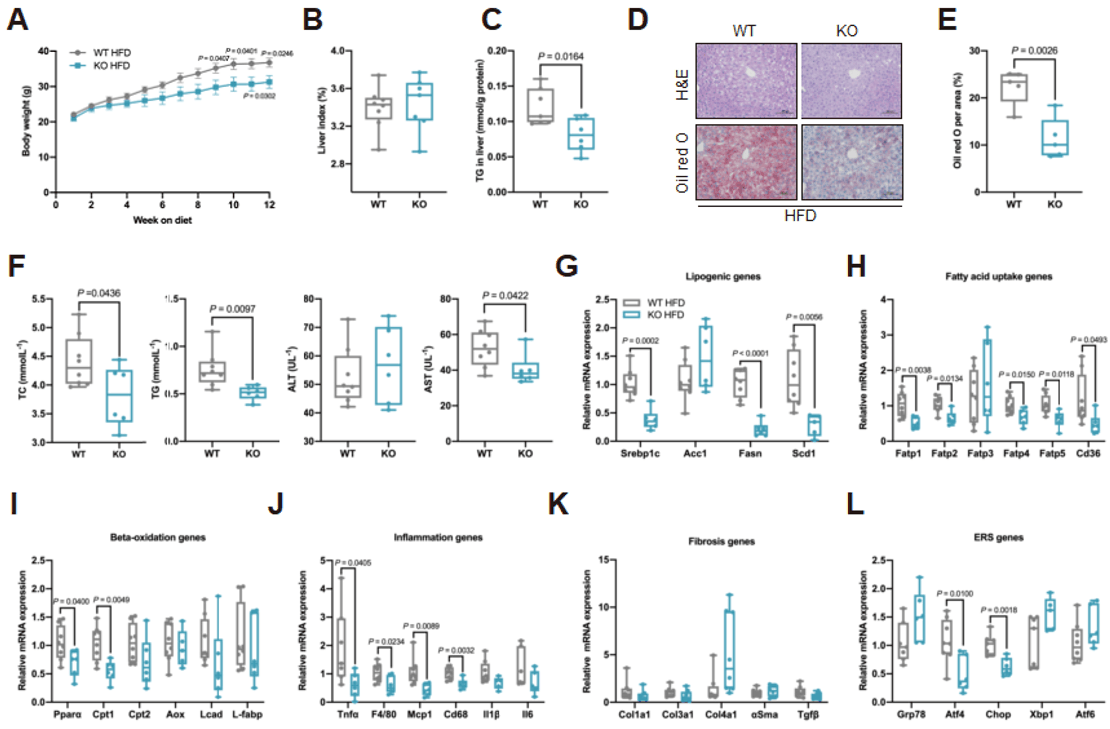
图1. HFD喂养背景下,Emc10 KO小鼠不易发生肝脂肪变性
EMC10消融加重MCD喂养小鼠的肝脂肪变性
EMC10对肝脂肪变性的影响在另一种脂肪变性动物模型中却有相反的结果。饲喂蛋氨酸-胆碱缺乏饮食(MCD)的小鼠常用作肝脏炎症和纤维化的脂肪变性模型,MCD喂养导致EMC10 KO组和WT组小鼠体重都明显减轻。与WT组相比,MCD喂养的EMC10 KO小鼠表现出更高的肝脏指数和更严重的肝脂肪变性,LFC、NAFLD活性评分(NAS)、血浆脂质和ALT(谷丙转氨酶)水平均较高;脂肪生成基因的表达显著减少,这可能是对甘油三酯分泌能力低下导致的过量LFC的负反馈反应;EMC10 KO小鼠肝脏中FA摄取基因的表达量高于WT小鼠,而β-氧化基因表达的显著降低;与加重的肝脂肪变性表型一致,EMC10 KO小鼠的肝脏炎症、纤维化和ER应激水平也有所增加。总之,与HFD喂养的模式相反,MCD喂养的EMC10 KO小鼠的肝脏脂肪变性加剧,结合上述结果表明EMC10以一种环境依赖的方式调节肝脂肪变性。
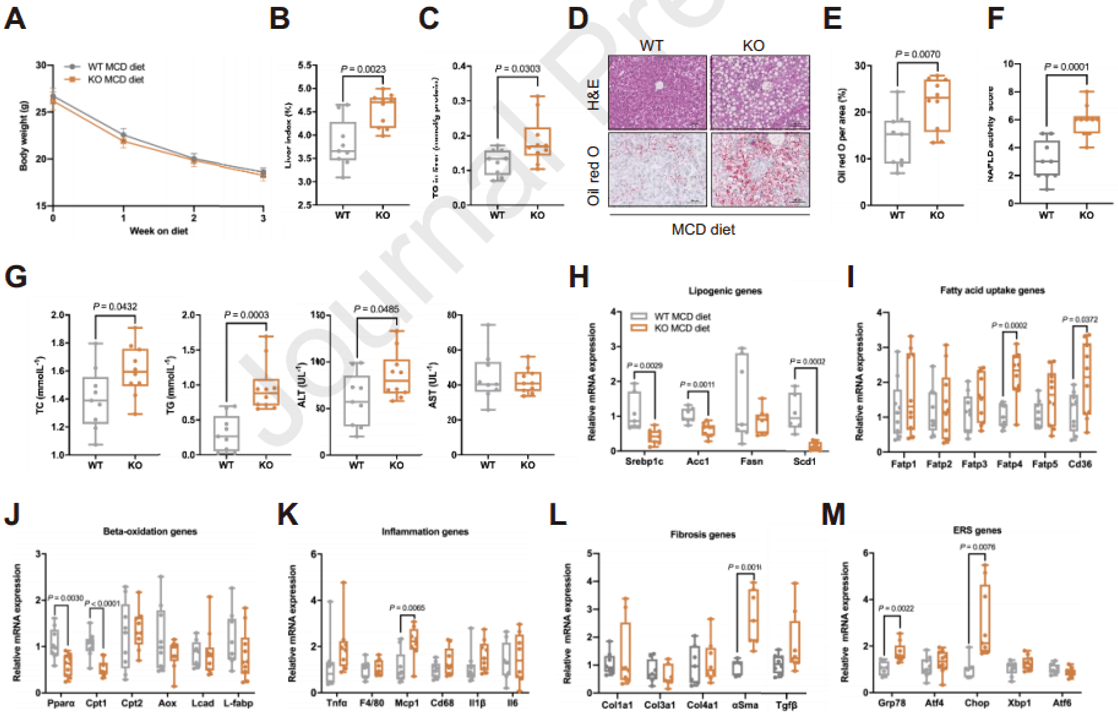
图2. MCD喂养条件下,Emc10 KO小鼠的肝脂肪变性加剧
在急性ER应激小鼠模型中,EMC10消融加重肝脂肪变性
由于EMC10 KO在小鼠肝脂肪变性的两种模型中的不同作用,促使作者进一步探索EMC10调节肝脂肪变性的潜在机制。前期实验已经发现HFD和MCD喂养都可以激活小鼠肝脏中的PERK-eIF2α信号传导,该信号通路是ER应激情况下,未折叠蛋白反应(UPR)的一个分支,被广泛认为与肝脂肪变性有关。作者通过衣霉素给药建立急性ER应激动物模型,衣霉素会显著诱导WT小鼠急性ER应激和因FA β氧化受损引起的肝脂肪变性。与对照相比,EMC10消融显著增加了衣霉素诱导的肝脂肪变性,KO小鼠的肝脏指数、LFC以及血清ALT、AST水平都高于WT型;并且KO小鼠肝脏的FA摄取显著增加,β氧化减少,纤维化加剧;Grp78、Atf4和Chop基因的表达也显著增加,而Xbp1和Atf6的表达没有变化,这说明UPR分支中的PERK-eIF2α通路被激活。结合MCD喂养小鼠中的结果,可以认为EMC10消融无法应对ER应激,进而通过激活肝脏PERK-eIF2α-ATF4信号加重了肝脂肪变性。
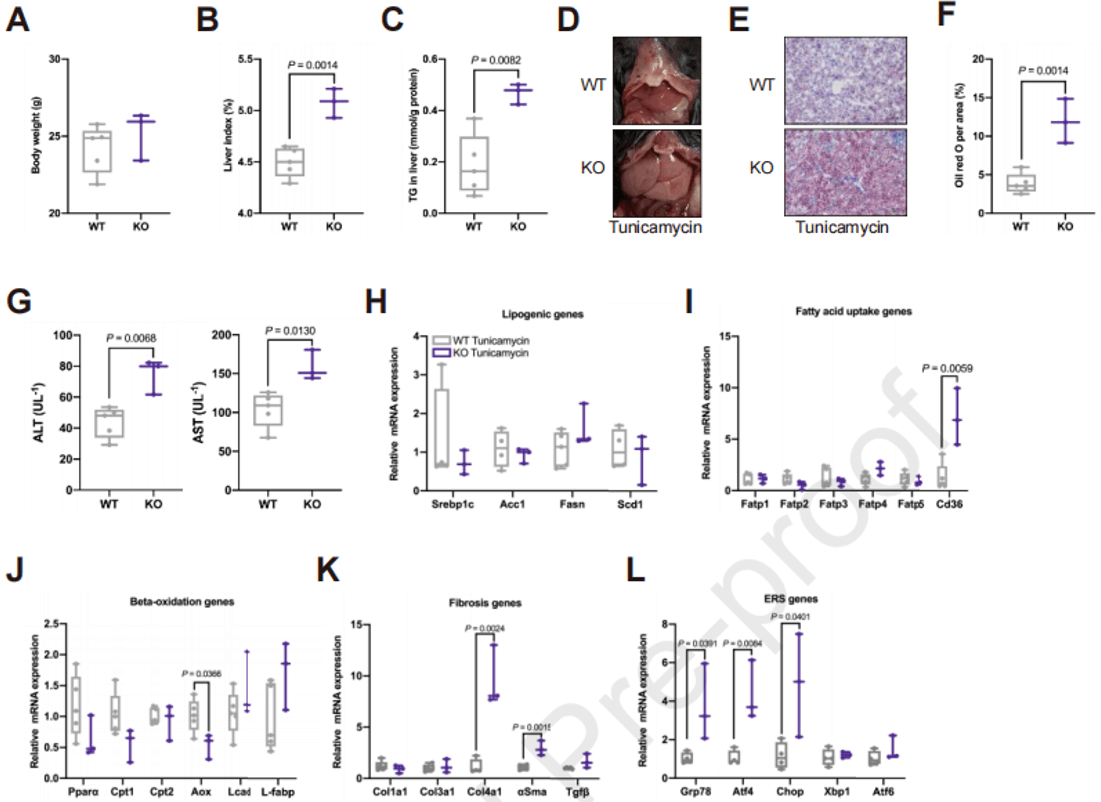
图3. 衣霉素引起Emc10 KO小鼠肝脂肪变性加重
EMC10调控小鼠肝脏中的PERK-eIF2α信号
为了阐明UPR在EMC10调节的肝脂肪变性中的作用,作者测定了EMC10 KO和WT小鼠肝脏中PERK-eIF2α的信号活性。在正常鼠粮喂养条件下,与WT对照组相比,EMC10 KO小鼠的PERK和eIF2α磷酸化水平以及肝脏中GRP78和CHOP蛋白丰度显著降低,说明信号传导被显著抑制;HFD饲喂后,EMC10 KO小鼠的eIF2α磷酸化水平也同样是显著降低。然而,当给予MCD饲喂或衣霉素处理时,与WT对照组相比,Emc10 KO小鼠肝脏的PERK-eIF2α-ATF4信号活性显著增加,这与它们加重的肝脂肪变性现象一致。这些数据表明,EMC10是维持肝脏UPR稳态所必需的。
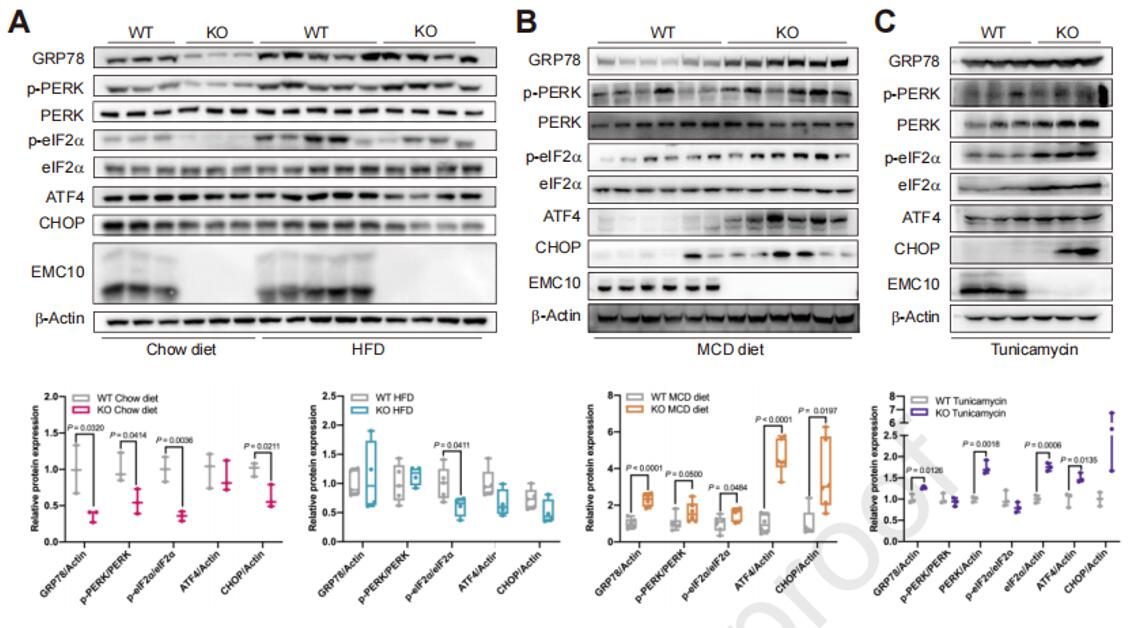
图4. EMC10调节肝脏PERK-eIF2α信号传导
EMC10的两种亚型在调节肝脏PERK-eIF2α信号传导方面存在差异
由于EMC10的两种亚型在一些细胞生理过程中具有不同的功能,所以作者探讨了这两种异构体是否以不同的方式调节肝脏PERK-eIF2α信号。在HepG2细胞中过表达mEMC10显著减轻了eIF2α信号以及细胞的脂质积累,说明mEMC10在肝细胞ER应激和脂肪变性中具有抑制作用;而scEMC10显著增强了HepG2细胞中的PERK-eIF2α信号传导活性,小鼠注射重组scEMC10也能显著增强肝脏中的PERK-eIF2α信号。当PERK的激活被化学抑制剂GSK2606414或siRNA抑制时,scEMC10诱导的eIF2α磷酸化增强会完全受损,说明scEMC10诱导的eIF2α激活依赖于PERK。
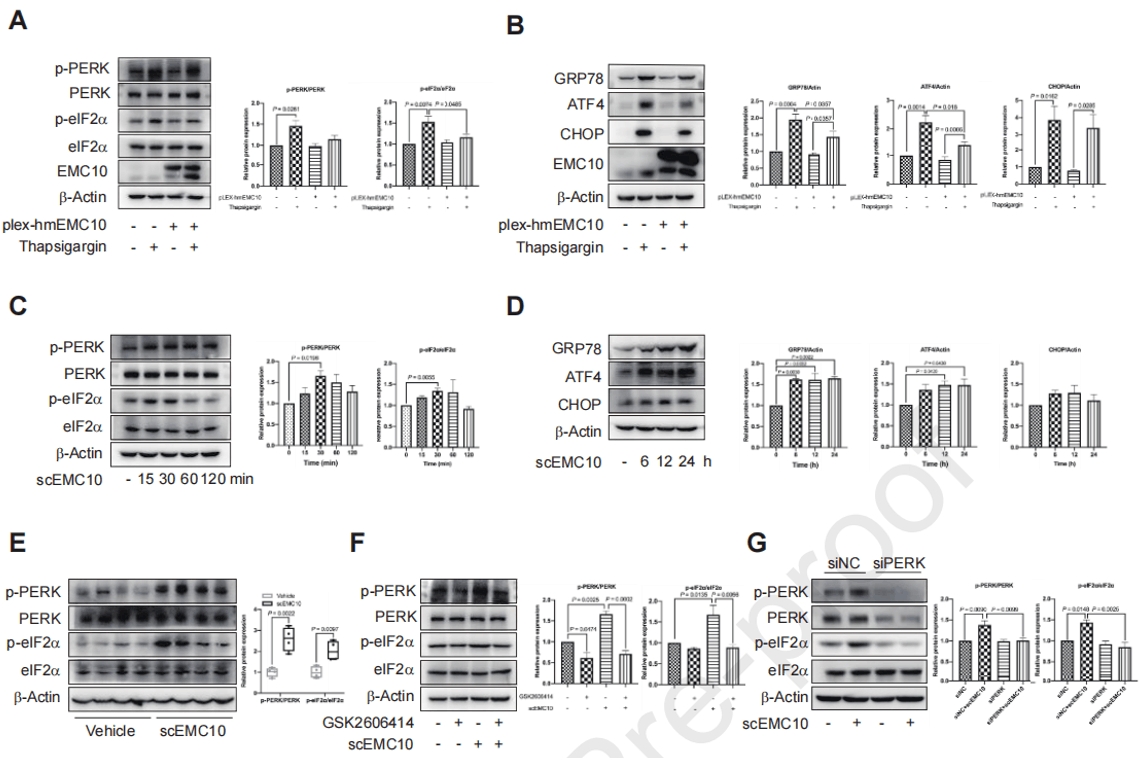
图5. mEMC10和scEMC10在肝脏PERK-eIF2α信号调节中的不同作用
scEMC10亚型促进小鼠肝脂肪变性的发展
在小鼠中利用AAV8过表达scEMC10,并给予HFD喂养,结果表明与注射AAV8空载对照相比,scEMC10的过表达使得小鼠的肝指数、LFC、NAS、血清ALT以及脂肪生成、FA摄取、炎症、纤维化和ER应激相关基因都显著升高,β-氧化基因表达降低,免疫印迹分析也表明scEMC10过表达的肝脏中eIF2α信号增强。这些结果表明scEMC10促进小鼠肝脂肪变性的发展。
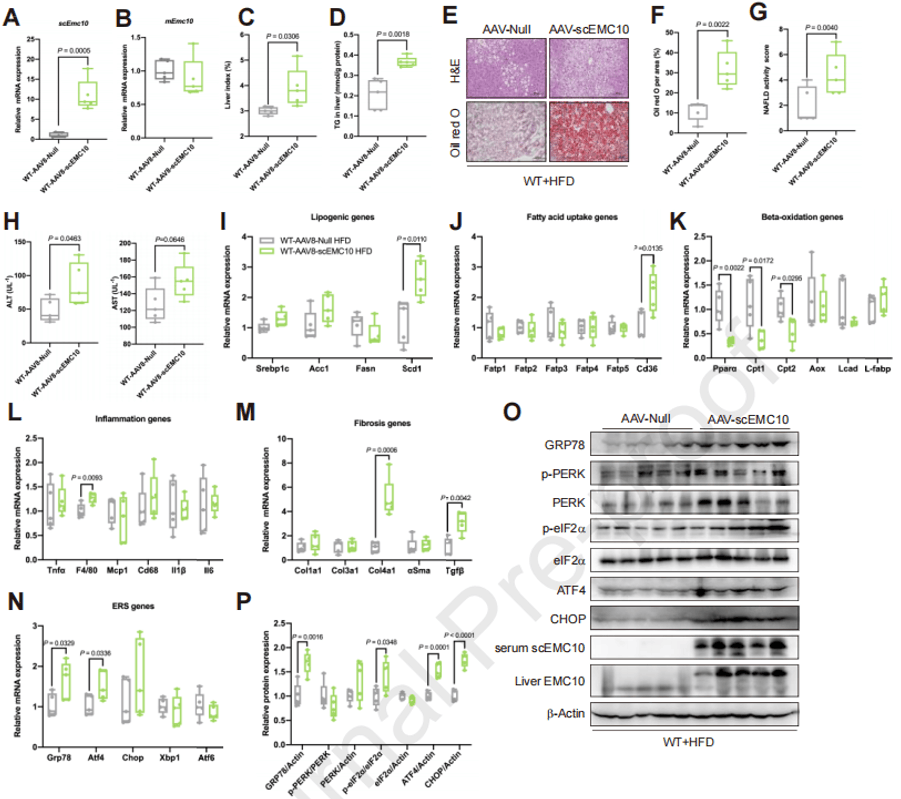
图6. 过表达scEMC10促进HFD喂养小鼠的肝脂肪变性
mEMC10亚型改善由衣霉素诱导的小鼠肝脂肪变性
作为一种循环因子,在肝脏中过表达的scEMC10可能靶向到达其他组织,这增加了scEMC10调节肝脂肪变性是间接的且不依赖于肝脏的可能性,所以作者又利用AAV8在小鼠肝脏中特异性过表达mEMC10。衣霉素诱导条件下,与对照组相比,mEMC10过表达小鼠的LFC和血清ALT、AST水平显著降低,β氧化增强,纤维化和ER应激减少,也就是说过表达mEMC10显著改善了衣霉素诱导的小鼠肝脂肪变性;并且蛋白质分析表明mEMC10对肝脏ER应激有显著的抑制作用。说明mEMC10在肝细胞减轻ER应激和肝脂肪变性ATF4活化中发挥直接作用。
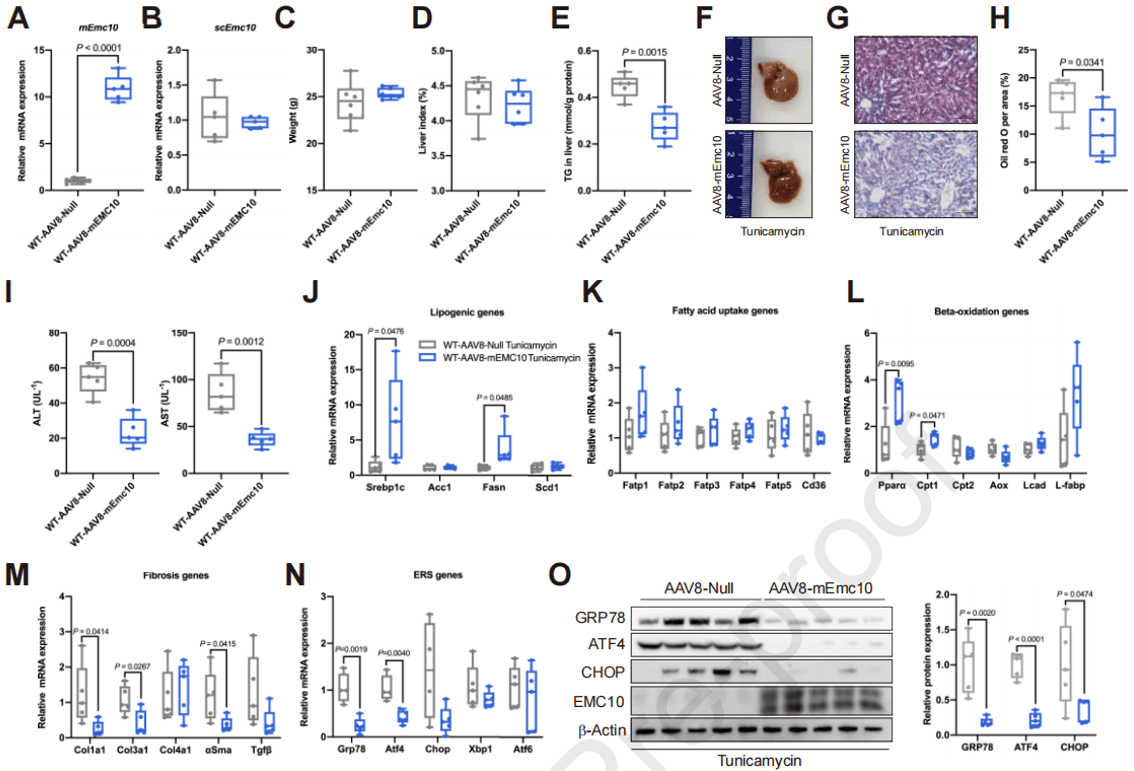
图7. 小鼠肝脏过表达mEMC10可改善衣霉素诱发的肝脂肪变性
中和循环的scEMC10可改善MCD喂养小鼠的肝脂肪变性
scEMC10中和抗体4C2(4C2 Ab)可以显著降低HFD喂养小鼠的肝脏脂质积累和血清ALT水平已经得到验证,那么在MCD饮食喂养中是否具有相同作用呢?与对照IgG相比,4C2 Ab导致小鼠肝脏指数、LFC、脂肪生成、FA摄取、炎症和ER应激水平均显著降低,而β氧化保持不变;MCD喂养小鼠肝脏中eIF2α和CHOP的磷酸化水平会增加,而4C2 Ab处理可以显著降低它们的磷酸化水平。这些发现表明scEMC10是治疗MASLD的一个潜力靶点。
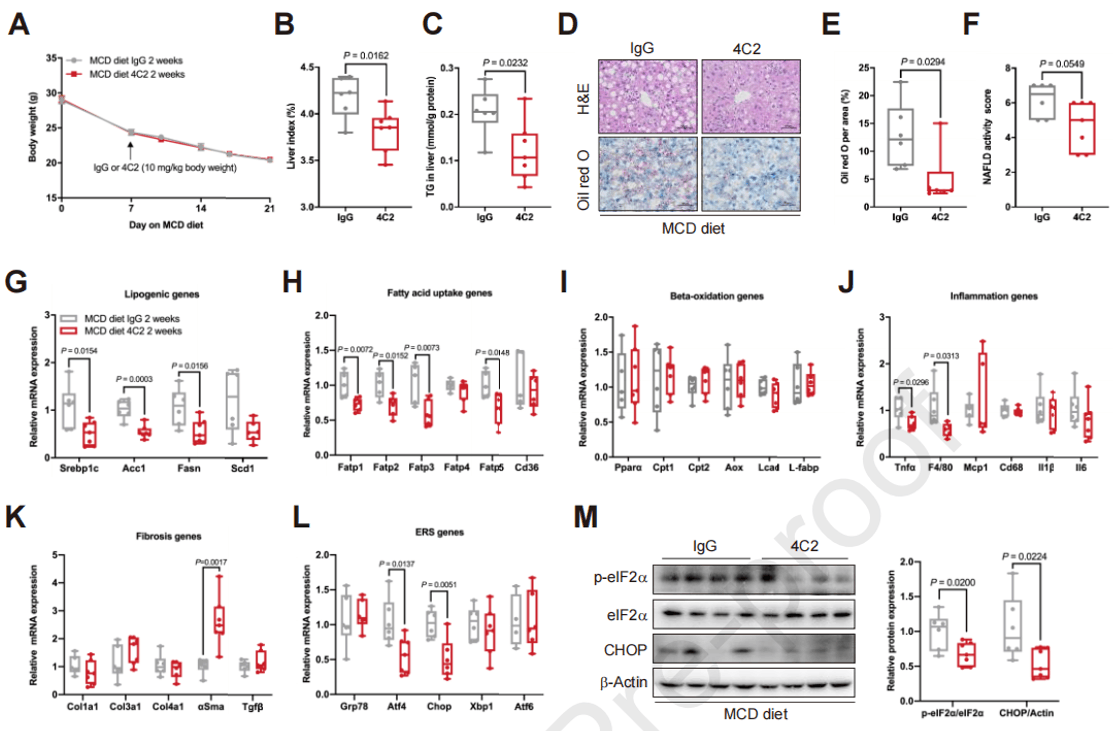
图8. 中和scEMC10可改善MCD喂养小鼠的肝脂肪变性
血清scEMC10与MASLD呈正相关,而肝脏mEMC10与MASLD呈负相关
为了探索scEMC10和mEMC10在人类MASLD中的作用,在高加索人的MASLD和非MASLD人群中测定了血清和肝脏scEMC10水平、LFC以及scEMC10、mEMC10、PERK-eIF2α信号下游靶基因的mRNA水平,观察到MASLD患者血清scEMC10水平随着LFC、NAS和纤维化评分的变化而显著升高;肝脏mEMC10 mRNA水平显著降低,但不同评分的MASLD中没有差异;表明血清scEMC10水平与LFC、肝scEMC10、ATF4和CHOP的mRNA水平以及血清ALT、AST和GGT(谷氨酰转移酶)呈正相关,肝mEMC10 mRNA水平与LFC、血清scEMC10、ALT、AST和GGT呈负相关。这些临床数据说明scEMC10与MASLD呈正相关,而肝脏mEMC10与MASLD呈负相关,也暗示mEMC10在抑制MASLD中发挥直接作用。
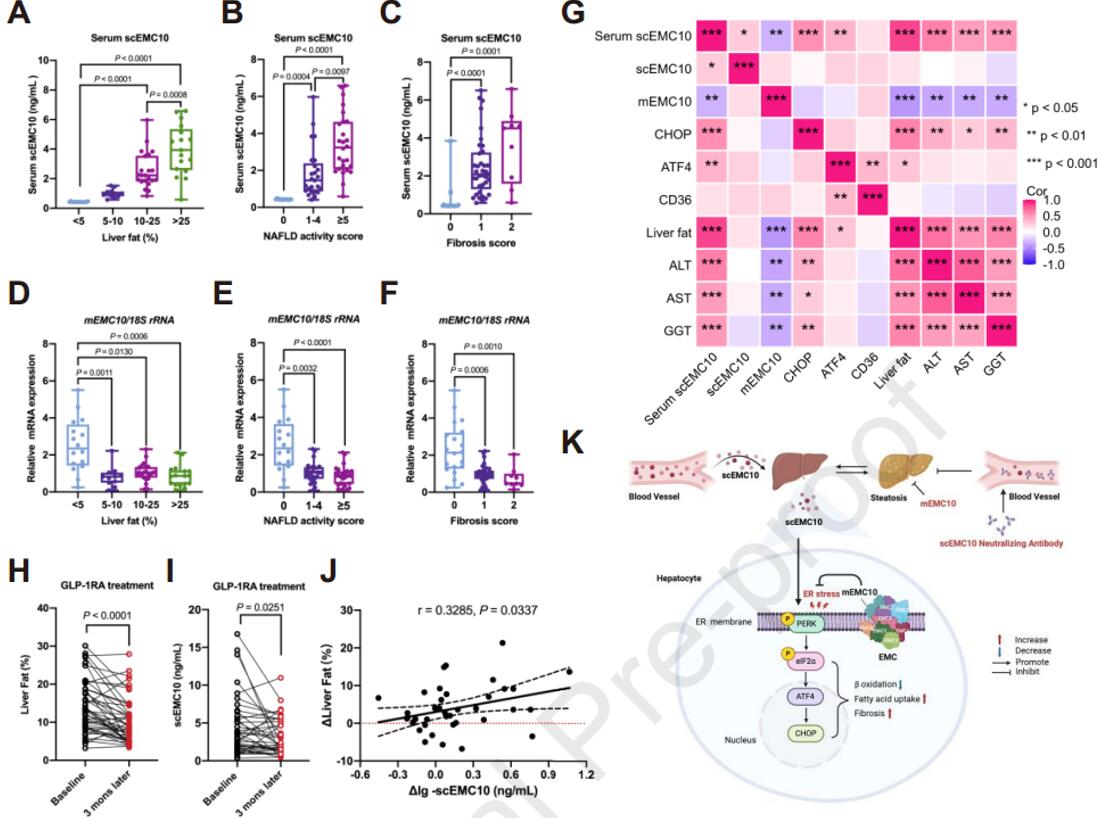
图9. EMC10与人类MASLD的关联
该研究表明,scEMC10促进小鼠肝细胞PERK-eIF2α-ATF4信号通路的激活以及与其相关的脂肪肝,而mEMC10则起抑制作用,并得到了临床研究结果的支持,血清scEMC10与人MASLD呈正相关,而肝脏mEMC10与之呈负相关。这项工作区分了EMC10的两种亚型在MASLD中的作用,并将scEMC10确定为MASLD的一个有前途的治疗靶点,为scEMC10调节治疗MASLD的转化提供了基础。









 3344
3344










